-
技術(shù)文章ARTICLE
CRISPR篩選和糖蛋白質(zhì)組學(xué)聯(lián)合研究發(fā)現(xiàn)新型WNT通路的N-糖基化調(diào)控機(jī)制
發(fā)布時(shí)間: 2025-11-13 點(diǎn)擊次數(shù): 90次——跟著Science學(xué)習(xí)如何進(jìn)行N-糖基化修飾的機(jī)制研究

美國(guó)斯坦福大學(xué)Rajat Rohatgi研究組發(fā)現(xiàn),受調(diào)控的N-糖基化控制伴侶蛋白功能和受體轉(zhuǎn)運(yùn)。這與將N-糖基化視為常規(guī)維護(hù)功能的普遍看法相反。研究人員通過(guò)N-糖肽組學(xué)分析揭示了內(nèi)質(zhì)網(wǎng)中一個(gè)調(diào)控N-糖基化的新途徑:該途徑由OST-A與HSP90B1(一種膜受體的ER伴侶蛋白)和CCDC134(一種ER腔內(nèi)蛋白)組成,在HSP90B1轉(zhuǎn)運(yùn)到ER的過(guò)程中,其N端肽段引導(dǎo)了包含CCDC134和OST-A的易位復(fù)合體的組裝,該復(fù)合體在折疊期間保護(hù)HSP90B1,防止其過(guò)度糖基化和降解。該通路對(duì)于WNT信號(hào)傳導(dǎo)至關(guān)重要,其功能障礙可能導(dǎo)致骨發(fā)育障礙。這一研究結(jié)果于2024年11月8日發(fā)表在Science雜志上,題為“Regulated N-glycosylation controls chaperone function and receptor trafficking"。
研究結(jié)果
基于CRISPR篩選激活WNT/β-catenin信號(hào)的基因
使用基于熒光的轉(zhuǎn)錄報(bào)告基因,在人類細(xì)胞系(RKO)中進(jìn)行了全基因組的功能缺失CRISPR/Cas9篩選,以鑒定響應(yīng)WNT3A激活WNT/β-catenin信號(hào)所需的基因。篩選發(fā)現(xiàn)了許多已知的WNT信號(hào)成分: β-catenin, LRP6 (WNT配體的共受體),以及兩種ER伴侶,MESD和HSP90B1,促進(jìn)LRP5和LRP6 (LRP5/6)在ER中的折疊。篩選還發(fā)現(xiàn)了三個(gè)與WNT信號(hào)不明確相關(guān)的基因:CCDC134、STT3A和OSTC。
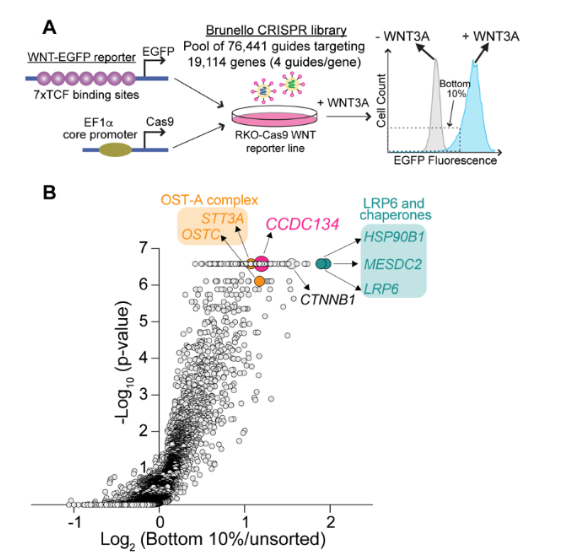
圖1. CRISPR/Cas9篩選鑒定鑒定響應(yīng)WNT3A激活WNT信號(hào)傳導(dǎo)的陽(yáng)性調(diào)節(jié)因子。
內(nèi)質(zhì)網(wǎng)伴侶HSP90B1的高度糖基化調(diào)控
敲除STT3A(而不是STT3B)導(dǎo)致HSP90B1的高度糖基化和去穩(wěn)定化。CCDC134和OSTC的敲除也導(dǎo)致HSP90B1的高度糖基化和不穩(wěn)定,此外,STT3A、CCDC134或OSTC的缺失降低了細(xì)胞表面WNT共受體LRP6的豐度。高度糖基化的HSP90B1不能折疊成功能蛋白,導(dǎo)致其被ERAD機(jī)制標(biāo)記和降解。N-糖蛋白學(xué)顯示STT3A和CCDC134與HSP90B1的N-糖基化具有高度特異性和一致性。

圖2. 內(nèi)質(zhì)網(wǎng)蛋白網(wǎng)絡(luò)對(duì)HSP90B1的N-糖基化和WNT信號(hào)的調(diào)控。
內(nèi)質(zhì)網(wǎng)中CCDC134對(duì)細(xì)胞表面WNT信號(hào)的調(diào)控
多種小鼠和人細(xì)胞系中CCDC134的敲除導(dǎo)致HSP90B1高度糖基化和不穩(wěn)定。回補(bǔ)實(shí)驗(yàn)表明,使用多xi環(huán)素誘導(dǎo)系統(tǒng)可以恢復(fù)CCDC134的表達(dá),抑制HSP90B1高度糖基化并恢復(fù)其豐度。此外,CCDC134的共表達(dá)抑制了其過(guò)表達(dá)引起的HSP90B1高度糖基化,并且在ER應(yīng)激下對(duì)HSP90B1具有劑量依賴性的保護(hù)作用。CCDC134-/-細(xì)胞表面LRP5/6豐度明顯降低。CCDC134-/-細(xì)胞對(duì)WNT配體的反應(yīng)遠(yuǎn)低于野生型細(xì)胞。CCDC134通過(guò)控制LRP5/6 (WNT配體的專性共受體)到細(xì)胞表面的運(yùn)輸來(lái)調(diào)節(jié)內(nèi)質(zhì)網(wǎng)中的WNT信號(hào)。CCDC134-/-患者的原代成纖維細(xì)胞表現(xiàn)出HSP90B1高度糖基化,細(xì)胞表面LRP6豐度降低,WNT信號(hào)通路受損,所有這些缺陷在CCDC134重新表達(dá)后被逆轉(zhuǎn)。

圖3. 在攜帶CCDC134突變的人類患者中 HSP90B1高度糖基化和LRP6。
CCDC134、OST-A和HSP90B1組成的信號(hào)通路
與CCDC134或HSP90B1的缺失類似,敲除STT3A(但不包括STT3B)也顯著降低了細(xì)胞表面LRP6和WNT信號(hào)的強(qiáng)度。值得注意的是,HSP90B15N的表達(dá)足以wan全挽救STT3A-/-細(xì)胞中的WNT信號(hào),盡管當(dāng)OST-A功能喪失時(shí),數(shù)百種膜和分泌蛋白的N-糖基化會(huì)發(fā)生改變。與CCDC134-/-細(xì)胞相比,STT3A-/-細(xì)胞中HSP90B1 N-糖基化有顯著差異。前者所有HSP90B1被高度糖基化,而后者只有一小部分被高度糖基化。即使CCDC134豐度大量增加,也無(wú)法抑制STT3A-/-細(xì)胞中的HSP90B1高度糖基化。這些數(shù)據(jù)表明,OST- B(STT3A-/-細(xì)胞中唯yi的OST復(fù)合物)在所有序列中默認(rèn)wan全N-糖基化HSP90B1,并且不能被CCDC134調(diào)節(jié)。在STT3B-/-細(xì)胞中,CCDC134的缺失導(dǎo)致HSP90B1高度糖基化,這表明CCDC134可以調(diào)節(jié)OST-A。在缺乏CCDC134的情況下,與OST-B相比,OST-A對(duì)HSP90B1的N-糖基化效率較低。
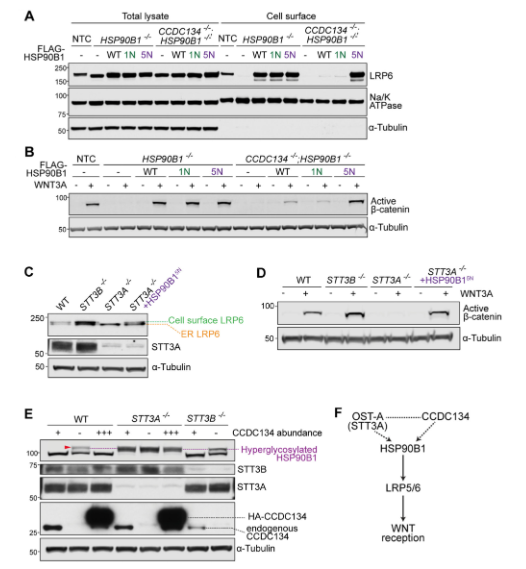
圖4. 高度糖基化的HSP90B1調(diào)控WNT信號(hào)通路
HSP90B1中的N端非結(jié)構(gòu)化肽調(diào)節(jié)其自身的N-糖基化修飾
系統(tǒng)缺失分析表明,HSP90B1的1-93氨基酸,包括ER信號(hào)序列和所有前N段,可以影響M結(jié)構(gòu)域遠(yuǎn)端序列的N-糖基化修飾。與STT3A-/-細(xì)胞中的HSP90B1相似,HSP90B1- T44A突變體也對(duì)CCDC134調(diào)控具有抗性。 HSP90B1的N端1~93個(gè)氨基酸,預(yù)計(jì)在很大程度上是非結(jié)構(gòu)化的,具有兩個(gè)自主的,可轉(zhuǎn)移的特性:它損害在同一多肽中緊隨其后的序列的N糖基化,也賦予CCDC134敏感性。HSP90B1的高度糖基化是由其自身N端、CCDC134和OST-A之間的復(fù)合物調(diào)節(jié)的模型,該復(fù)合物在其翻譯和轉(zhuǎn)運(yùn)到內(nèi)質(zhì)網(wǎng)時(shí)組裝。
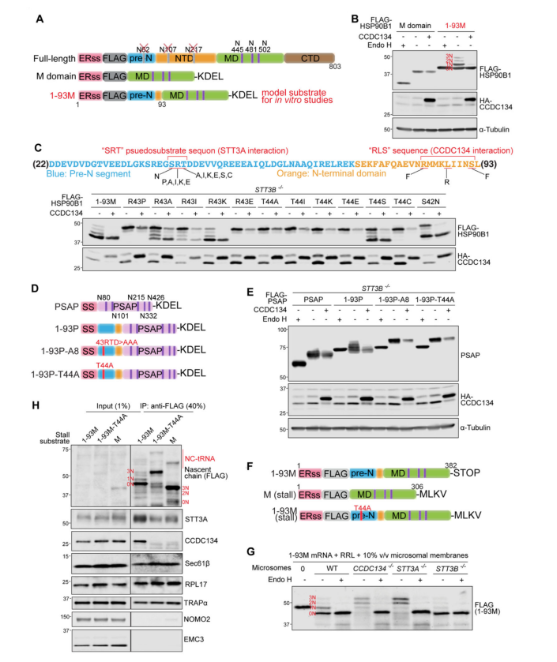
圖5. HSP90B1的前體N段通過(guò)招募CCDC134到含有OST-A的易位子來(lái)抑制其自身的N-糖基化。
OST-A作用機(jī)制
OST-A的糖基轉(zhuǎn)移酶活性對(duì)于抑制HSP90B1過(guò)度糖基化并非必需。這一發(fā)現(xiàn)挑戰(zhàn)了我們對(duì)OST-A功能的傳統(tǒng)理解,提示OST-A可能通過(guò)非酶活性的方式參與蛋白質(zhì)的折疊和質(zhì)量控制。研究人員通過(guò)構(gòu)建OST-A的突變體,并在細(xì)胞中表達(dá)這些突變體,發(fā)現(xiàn)即使在OST-A的糖基轉(zhuǎn)移酶活性被抑制的情況下,HSP90B1的糖基化仍然受到抑制,表明OST-A可能通過(guò)其他機(jī)制調(diào)控HSP90B1的糖基化。OST-A可能不是作為一種酶起作用,而是作為一種支架起作用。

圖6. HSP90B1的穩(wěn)定性和WNT信號(hào)傳導(dǎo)不依賴OST-A的寡糖轉(zhuǎn)移酶活性。
文章總結(jié)
一、主要方法
· 利用細(xì)胞生物學(xué)方法(如免疫印跡、共沉淀、顯微鏡定位)分析蛋白修飾與相互作用。
· 通過(guò)突變體(改變N-位點(diǎn)或糖基化酶活性)、化學(xué)抑制劑(如抑制糖鏈修剪或加成)調(diào)控糖基化過(guò)程。
· 功能性測(cè)定包括受體在細(xì)胞表面的表達(dá)量、配體結(jié)合/信號(hào)轉(zhuǎn)導(dǎo)能力及降解速率等。
· 質(zhì)譜分析糖基化位點(diǎn)、以及生化方法分析糖鏈結(jié)構(gòu)變化。
二、核心發(fā)現(xiàn)
1. N-糖基化是可調(diào)控的,并且其狀態(tài)(是否附有完整糖鏈或是否已被糖苷酶/糖轉(zhuǎn)移酶修飾)決定了伴侶蛋白對(duì)底物的識(shí)別與結(jié)合強(qiáng)度。
2. 伴侶蛋白(如calnexin/calreticulin)對(duì)特定糖基化狀態(tài)的偏好影響其與新生受體蛋白的相互作用時(shí)間,從而改變折疊效率與質(zhì)量控制決策(繼續(xù)折疊/進(jìn)入降解通路)。
3. 改變N-糖基化(通過(guò)基因突變或化學(xué)干預(yù))會(huì)直接影響某些受體(可能為膜受體或分泌受體)的成熟與從ER到高爾基體及細(xì)胞膜的轉(zhuǎn)運(yùn),導(dǎo)致細(xì)胞表面受體量和功能發(fā)生變化。
4. 該調(diào)控機(jī)制在細(xì)胞對(duì)壓力或生理信號(hào)響應(yīng)時(shí)可能被動(dòng)員,以調(diào)整細(xì)胞表面受體數(shù)量或調(diào)節(jié)分泌蛋白的輸出,從而參與更廣泛的細(xì)胞功能與適應(yīng)性反應(yīng)。
三、結(jié)論與意義
· N-連接糖基化不僅是結(jié)構(gòu)性修飾,也是動(dòng)態(tài)調(diào)控蛋白命運(yùn)的關(guān)鍵信號(hào)。
· 通過(guò)控制伴侶蛋白與底物的相互作用,糖基化狀態(tài)決定受體是否通過(guò)質(zhì)量控制并正確定位到細(xì)胞表面。
· 理解這個(gè)調(diào)控軸有助于解釋某些遺傳病、代謝或分泌失調(diào)的分子機(jī)制,并為開發(fā)針對(duì)糖基化或伴侶系統(tǒng)的小分子干預(yù)提供理論依據(jù)。
四、后續(xù)研究方向
· 識(shí)別更廣泛的受體/底物譜及其敏感的糖基化位點(diǎn)。
· 解析不同細(xì)胞類型或生理狀態(tài)下糖基化調(diào)控的機(jī)制(例如糖酶、轉(zhuǎn)移酶的調(diào)控)。
· 開發(fā)藥物以修復(fù)因糖基化異常導(dǎo)致的蛋白折疊或定位缺陷。

產(chǎn)品中心
Products
![]()
技術(shù)支持:環(huán)保在線 地址:上海市寶山區(qū)長(zhǎng)江南路180號(hào)B區(qū)B650 管理登陸 sitemap.xml
 微信二維碼
微信二維碼擊這里給我發(fā)消息")
